Le chou lève le voile sur ses fractales
L’origine de la forme fractale du chou-fleur et du chou romanesco a enfin été élucidée ! Un article publié dans la revue « Science » détaille les contrôles génétiques à l’origine de cette structure si particulière. Un dérèglement moléculaire empêche les bourgeons de fleurir et entraine leur multiplication infinie. Une fleurette initiale forme alors un ensemble de fleurettes formant chacune un ensemble de fleurettes…
La structure de la pomme des choux
Parmi la diversité des choux (Brassica oleracea), le chou-fleur, le brocoli et le chou romanesco ont la particularité de former des inflorescences.
En effet, la partie comestible de ces choux est un organe préfloral hypertrophié, appelé pomme, que l’on pourrait comparer à un regroupement de petits bourgeons organisés en spirales et formant des fleurettes.
La pomme du chou-fleur, comme celle du chou romanesco, présente la particularité de posséder une structure fractale, c’est-à-dire que l’on retrouve une structure similaire (fleurette) sur l’ensemble de la pomme et à toutes les échelles : chaque fleurette est constituée de spirales faites de fleurettes plus petites et ainsi de suite.
Des mutations génétiques sont à l’origine de la structure fractale des pommes de choux. La formation d’une telle structure a longtemps intrigué les chercheurs, aussi ont-ils réuni des compétences à la fois de modélisation mathématique et de biologie végétale pour élucider cette énigme.
Le contrôle génétique du développement des bourgeons
Une plante développe ses différents organes à partir des bourgeons. Ceux-ci produisent une tige présentant elle-même des bourgeons. Ces nouveaux bourgeons sont en général dans un état dormant, laissant la tige croitre, puis ils sont activés pour former à leur tour de nouvelles tiges. Lorsque les conditions environnementales et le développement de la plante sont favorables à la floraison, un signal moléculaire réoriente les bourgeons vers un développement floral. Le passage du bourgeon de l’état « tige » à l’état « fleur » est irréversible, ce dernier sera alors amené à produire une fleur ou une inflorescence.
L’attribution de l’identité du bourgeon s’opère via une « guerre de territoire » telle que l’a décrite François Parcy dans son livre « l’histoire secrète des fleurs ». Nous avons d’un côté un « gardien de la tige », le gène TFL1 (Terminal Flower 1) et de l’autre côté « l’architecte floral », le gène LFY (LEAFY). Le gardien de la tige inhibe l’architecte floral pour maintenir l’état « tige » du bourgeon. Cependant lorsque les conditions sont favorables, d’autres facteurs interviennent pour activer le gène LFY. Une fois activé, l’architecte floral active à son tour d’autres gènes inhibant le gène TFL1 (« gardien de la tige »), empêchant alors le retour du bourgeon à l’état « tige ». Parmi ces gènes, nous trouvons AP1 et CAL. Une cascade d’expression de gènes aboutit alors à la formation des fleurs.
Les chercheurs ont construit un modèle représentant les interactions moléculaires contrôlant l’identité du bourgeon et donc la floraison. Ce modèle leur a permis d’identifier les mutations responsables de la formation de la pomme du chou-fleur.
Les mutations à l’origine de la formation d’une pomme de chou-fleur
Pour comprendre les mécanismes génétiques qui contrôlent la formation de la pomme du chou-fleur, une espèce modèle a été utilisée, il s’agit de l’arabette des dames (Arabidopsis thaliana). Cette espèce est très étudiée en raison de son génome de petite taille (elle présente 5 paires de chromosomes contre 9 chez le chou-fleur). Cela permet d’étudier plus facilement son génome à la recherche de mutations pouvant expliquer des différences observables à l’échelle de la plante. De plus, il s’agit d’une petite plante dont le cycle de développement complet (de graine à graine) est de seulement 6 semaines. Chez ce modèle, il a été observé que la mutation simultanée de 2 gènes entrainait la formation d’une structure semblable à un chou-fleur ! (Figure 1). Il s’agit des gènes AP1 et CAL (Azpeitia et al., 2021).

Figure 1: structure semblable au chou-fleur chez Arabidopsis thaliana (à droite).
Godin et Parcy, 2021.
Arabidopsis thaliana, est une espèce apparentée aux choux, elle appartient au même genre : les Brassica (anciennement appelées Crucifères). Les gènes identifiés chez cette espèce modèle ont été retrouvés par homologie de séquences chez le chou-fleur et les 2 mutations affectant ces gènes ont également été observées chez ce dernier.
Les mutations des gènes AP1 et CAL les rendent non fonctionnels. Ainsi, lorsque le bourgeon se transforme en bourgeon floral (activation du gène LFY), le gène TFL1 n’est pas inhibé par AP1 et CAL comme il devrait, et le bourgeon retourne à l’état « tige ». Cependant le passage à l’état floral étant irréversible, le retour à l’état « tige » n’est pas complet. Le bourgeon se met à croître immédiatement et reproduit des bourgeons floraux qui n’aboutissent pas non plus à une fleur mais redeviennent des bourgeons de tige, qui, à leur tour vont reformer des bourgeons floraux et ainsi de suite. Les bourgeons se multiplient formant une imbrication continue de tiges construisant la forme fractale si particulière du chou-fleur.
La figure 2 décrit les différents mécanismes génétiques conduisant à la formation de tiges, de fleurs ou de pommes.
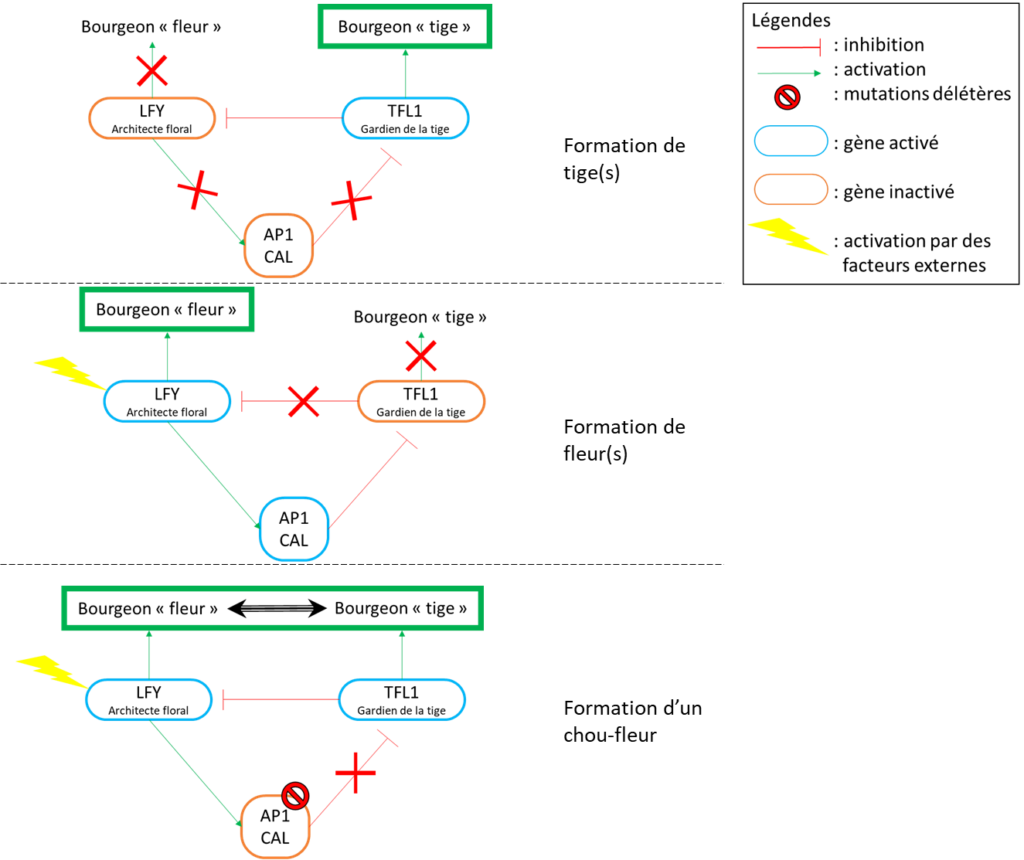
Figure 2: contrôle génétique du développement des bourgeons
Chez le chou romanesco, c’est l’aspect pyramidal des fleurettes, qui lui est propre, qui accentue et révèle sa structure fractale hypnotisante et fascinante. Une étude a été menée en 1998 par Kieffer et al. pour expliquer cette différence. La forme pyramidale des fleurettes du Romanesco serait due à une accélération du rythme de production des bourgeons qui est stable chez le chou-fleur. Cependant, les gènes à l’origine de ce phénomène n’ont pas encore été identifiés.
Conclusion :
L’article publié dans Science apporte un éclairage sur les interactions moléculaires complexes contrôlant la floraison et aboutissant à la forme si particulière des pommes de chou-fleur et de chou romanesco. Ces travaux de recherche ouvrent des perspectives intéressantes en agronomie et en sélection variétale, basées sur l’exploitation de ces mécanismes moléculaires.
Sources
Azpeitia, Eugenio, Gabrielle Tichtinsky, Marie Le Masson, Antonio Serrano-Mislata, Jérémy Lucas, Veronica Gregis, Carlos Gimenez, et al. « Cauliflower Fractal Forms Arise from Perturbations of Floral Gene Networks ». Science 373, nᵒ 6551 (9 juillet 2021): 192‑97. https://doi.org/10.1126/science.abg5999.
Godin, Christophe, et Francois Parcy. « Le Voile Se Lève Sur La Forme Fractale Du Chou-Fleur et Du Romanesco ». The Conversation. Consulté le 6 août 2021. http://theconversation.com/le-voile-se-leve-sur-la-forme-fractale-du-chou-fleur-et-du-romanesco-164730.
Parcy F. L’Histoire secrète des fleurs, HuenSciences, 240p (2019)
Kieffer, Martin, Michael P. Fuller, et Anita J. Jellings. « Explaining Curd and Spear Geometry in Broccoli, Cauliflower and `romanesco’: Quantitative Variation in Activity of Primary Meristems ». Planta 206, nᵒ 1 (1 juillet 1998): 34‑43. https://doi.org/10.1007/s004250050371.
Azpeitia, Eugenio, Gabrielle Tichtinsky, Marie Le Masson, Antonio Serrano-Mislata, Veronica Gregis, Carlos Gimenez, Nathanaël Prunet, et al. « The Making of Cauliflowers: The Story of Unsuccessful Flowers ». Report or Paper, 13 février 2021. https://resolver.caltech.edu/CaltechAUTHORS:20210216-122056927.
